


















































The Quest for Large-Scale DNA
Although genome editing was not a new concept, as zinc finger and TALEN platforms were already in use, the discovery of CRISPR-Cas9 shifted genome-editing research and clinical translation into high gear. But just like other platforms, this new kid on the block was not applicable to every editing situation for every genetic disease.
A longstanding desire in the field is a one-and-done, mutation-agnostic cure for genetic diseases that result from numerous mutations in a gene or from large-scale chromosomal structural variations, including deletions, duplications, inversions, and translocations. Gene therapies for these genetic indications require large-scale DNA manipulation, presenting different technical and regulatory challenges than correcting single-nucleotide point mutations.
Scientists donned their Indiana Jones hats to search for this holy grail of genome editing. Promising approaches under exploration included bridge recombinases, large serine recombinases, and CRISPR-associated transposases (CASTs), as well as immune-evasive DNA cargoes like circular single-stranded DNA (cssDNA), which may address the innate toxicity of double-stranded DNA (dsDNA) payloads.
Still, delivery can remain a conundrum for large payloads. For the most part, current delivery mechanisms are size-limited in terms of payloads, as are the workarounds using mRNA formats and leveraging reverse transcriptase.
The thirst is there, and the quest will continue. New genome-editing tools applicable to large DNA cargoes and delivery mechanisms will be refined, putting potential cures in sight for some deplorable diseases.
Bridge recombinases
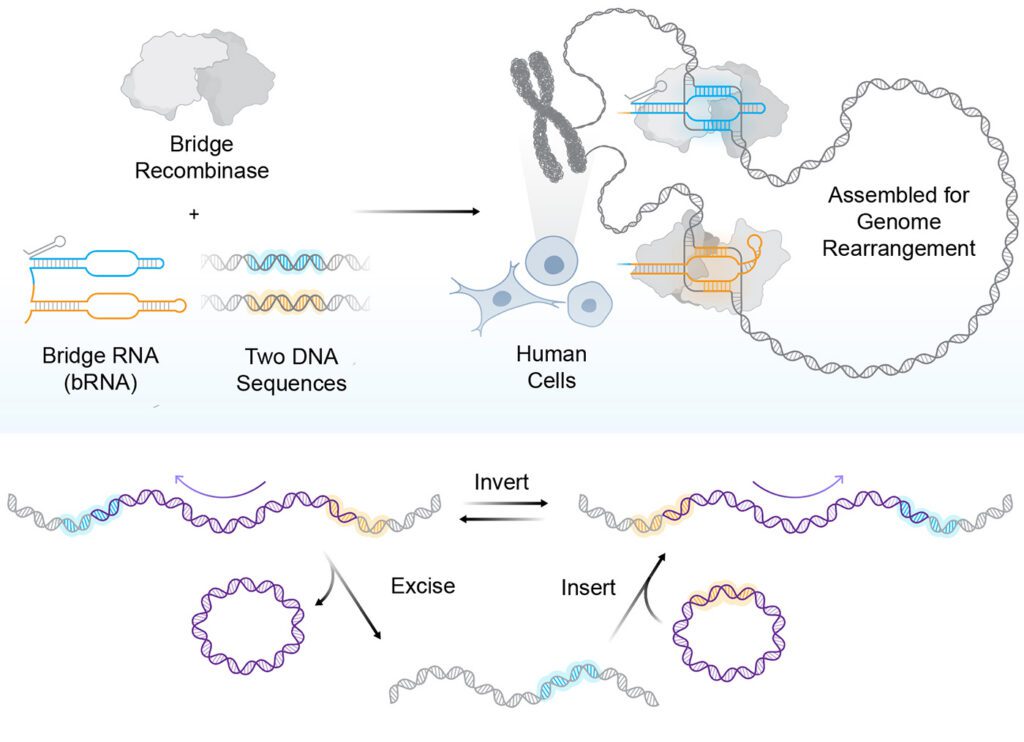
A new class of programmable genome-editing tools, bridge recombinases are the first RNA-guided DNA recombinases providing a distinct mechanism for manipulating DNA.1,2
The system has two key components: the recombinase enzyme, which catalyzes the DNA rearrangement, and a bridge RNA guide with two independently programmable loops. The target-binding loop controls genomic locus targeting, and the donor-binding loop specifies the donor payload.
Reprogramming the bridge RNA to change the configuration and orientation of the target and donor sites allows the system to be redirected to perform excision of a desired sequence from the genome or inversion of a DNA segment in place. The modularity means a single two-component system can perform all three fundamental DNA rearrangements—insertion, excision, and inversion—through a single unified mechanism.
“The most immediate advantage is the scale of DNA that bridge recombinases can manipulate,” said Patrick Hsu, PhD, co-founder and core investigator of the Arc Institute and assistant professor of pathology at Stanford University. “A technology that can operate at the scale of whole-gene replacement or correct structural variants opens up a new class of genetic interventions.” Multi-kilobase insertions, inversions up to 0.93 Mb, and excisions up to 0.13 Mb have all been demonstrated in human cells.3
Bridge recombination also does not rely on dsDNA breaks. The recombinase catalyzes strand exchange directly through a covalent intermediate, making the outcome deterministic in a way that nuclease-dependent approaches are not. This indicates that the system may have advantages in post-mitotic cells for therapeutic applications. About a quarter of the size of Cas9, the system can be encoded in delivery vectors with limited capacity.
“The technology is still in development. While our current efficiency and specificity numbers (20% insertion efficiency with 82% on-target) represent a meaningful proof-of-concept in human cells, improving both metrics will be necessary for safe and effective therapeutic applications,” said Hsu. The diversity of bridge-recombinase systems found in nature continues to be explored.
Large serine recombinases
“For simpler cases where we want to insert a DNA payload into a fixed safe harbor site, we are working on large serine recombinases (LSRs),” said Hsu. While these enzymes lack the RNA programmability of bridge recombinases, they offer very high efficiency and specificity of insertion and are effectively unidirectional, leading to very stable insertions of large DNA cargoes into the human genome.
Stylus Medicine, a company Hsu co-founded, intends to advance LSRs for in vivo genetic therapies. “I am excited to see the new therapies that will emerge from combining recombinase technology with machine learning-assisted protein engineering and advances in DNA and effector delivery for challenging disease contexts,” said Hsu.
CRISPR-associated transposases
CASTs are naturally occurring bacterial systems that utilize nuclease-deficient CRISPR machinery to integrate DNA at genomic locations specified by guide RNAs (gRNA). “While CRISPR is often used to cut DNA, CASTs instead use CRISPR systems to guide site-specific DNA transposition,” said Isaac Witte, PhD, department of chemistry and chemical biology at Harvard University.
In 2019, two research groups—one at Columbia University led by Sam Sternberg, PhD, and the other at the Broad Institute of MIT and Harvard, headed by Feng Zhang, PhD—found that CASTs use CRISPR systems to target DNA transposition by a transposase complex. Further work demonstrated that CASTs were very efficient bacterial genome editors.
CASTs can mobilize multi-kilobase-scale DNA cargoes, and their naturally evolved transposition mechanism avoids forming dsDNA breaks in the genome. The problem was that the wild-type systems exhibited extremely low (often ≤0.1% of treated cells) or undetected integration activity in human cells.
Collaborating with the Sternberg lab, the lab of David Liu, PhD, from the Broad Institute of MIT and Harvard, used PACE (phage-assisted continuous evolution), a directed evolution platform developed by the Liu lab, to enhance the efficiency of CAST transposition.
In PACE, bacteriophages, which infect host bacteria, encode evolving genes in place of an essential gene for phage replication. This essential gene is instead encoded by host bacteria. “In PACE, you link the desired activity of the evolving biomolecule to the expression of this essential gene. In this case, we linked targeted DNA integration to the replication of phages encoding evolving CAST protein components,” said Witte.
A series of modifications ensured efficient enhancement of activity, resulting in the generation of an evolved variant of the CAST transposase protein TnsB that mediated over 200-fold improved integration activity in human cells. The TnsB protein contained ten individual mutations scattered throughout the predicted structure, which contributed to improved activity.
The evolved TnsB was combined with other PACE-evolved and rationally engineered CAST components to yield evoCAST, a system optimized for human-cell integration activity, published in Science.5
The evoCAST DNA integration does not require formation of dsDNA breaks in the genome, resulting in undetected levels of insertion and deletion mutations (indels) commonly found in traditional methods of gene insertion like nuclease-stimulated, homology-directed repair (HDR). In addition, evoCAST can be easily reprogrammed to genomic sites of interest by changing the gRNA sequence, and it supports a variety of DNA payload sizes, ranging from less than 1 kb to at least 15 kb.
A potential limitation, however, is that evoCAST is molecularly complex, containing seven distinct protein subunits, making the total coding size (~8.5kb) relatively large compared to around 5 kb for Cas9.
Big-picture limitations center on delivery, according to Witte, such as mitigating the cytotoxicity of foreign dsDNA in most therapeutically relevant cell types. Additionally, reducing the size and the number of distinct components required for integration activity may facilitate evoCAST applications in vivo. Next steps include harnessing the naturally existing diversity of CAST systems to develop a more diverse repertoire of CASTs for genome editing in human cells.
Circular single stranded DNA
Full Circle Therapeutics’ genome writing technology centers on an immune evasive DNA modality, a mini-cssDNA, called C4DNA—circular, clean, concealed, and customizable up to 20 kb.
The company has commercialized over 350 research-grade cssDNA for primary sequences and is developing processes for GMP-grade products for clinical applications. According to Wu, initially, the company’s founder, Richard Shan, intended to supply linear cssDNA as a DNA commodity for researchers in the gene-integration field. The starting material was cssDNA that was cleaved into linear strands. A serendipitous benchmark experiment using cssDNA as a control demonstrated surprisingly better integration performance than its linear counterpart.6 The unexpected results led to a foundational patent describing the use of cssDNA for targeted genomic integration.
After benchmarking the different DNA formats, cssDNA appeared superior and compatible with various CRISPR-Cas systems, along with other meganuclease editing systems such as TALEN. Next, they evaluated the hypothesis that immunogenicity due to dsDNA could be eliminated if mobile genetic elements like transposase systems and LSRs could use cssDNA.
Collaborating with a team at Harvard Medical School led by Benjamin Kleinstiver, PhD, they demonstrated that naked unmodified cssDNA, combined with piggyback transposases or LSRs, enables kilobase writing, albeit inefficiently. One way to improve integration efficiency was to design an oligo that could fuse to the cssDNA with hydrogen bonding to form a partial duplex. A 30- to 60-mer partial duplex showed good integration efficiency when compared to dsDNA, while remaining immune silent.
In another approach, the team modified the nuclear editor and installed a peptide sequence identified from a bacterial genome with a strong binding affinity with cssDNA. In this case, the modified Cas9 became an engineered molecular chaperone to recruit the DNA molecule and form a complex, effectively loading and delivering the genome engineering complex into the nucleus and direct to the targeted genome.7
Continued collaboration with Kleinstiver’s lab aimed to improve integration efficiency. The approach, in this case, used a partial duplex cssDNA that reconstituted a recombinase recognition sequence. The scientists termed this integration through nucleus-synthesized template addition of large lengths (INSTALL). INSTALL is compatible with diverse genome engineering nucleases and RNA-guided recombinases for high-fidelity kilobase-scale human genome writing.8
“We welcome partners,” said Wu. “It is prime time to talk about DNA medicines.”
References
- Hiraizumi, M, Perry NT, Durrant, MG, et al. Structural mechanism of bridge RNA-guided recombination. Nature2024; 630:994-1002. doi:10.1038/s41586-024-07570-2
- Durrant, MG, Perry NT, Pai JJ, et al. Bridge RNAs direct programmable recombination of target and donor DNA. Nature2025:630:984-993. doi:10.1038/s41586-024-07552-4
- Perry NT, Bartie LJ, Katrekar D, et al. Megabase-scale human genome rearrangement with programmable bridge recombinases. Science. 2026 Mar 12;391(6790):eadz0276. doi:10.1126/science.adz0276
- Fanton, A, Bartie, LJ, Martins JQ, et al. Site-specific DNA insertion into the human genome with engineered recombinases. Nat Biotechnol. 2025 Nov 6. doi:10.1038/s41587-025-02895-3
- Witte IP, Lampe GR, Eitzinger S, et al. Programmable gene insertion in human cells with a laboratory-evolved CRISPR-assoc iated transposase. Science. 2025 May 15;388(6748). doi:1126/science.adt5199
- Xie K, Starzyk J, Majumdar I, et al. Efficient non-viral immune cell engineering using circular single-stranded DNA-mediated genomic integration. Nat Biotechnol. 2025 Nov;43(11):1821-1832. doi:10.1038/s41587-024-02504-9
- Nam H, Xie K, Majumdar I, et al. Engineering tripartite gene editing machinery for highly efficient non-viral targeted genome integration. Nat Commun. 2025; 16:4569. doi:1038/s41467-025-59790-3
- Tou CJ, Xie K, Ferreira da Silva J, et al. Immune evasive DNA donors and recombinases license kilobase-scale writing. Nature. 2026 Mar 11. doi:10.1038/s41586-026-10241-z
The post The Quest for Large-Scale DNA appeared first on GEN - Genetic Engineering and Biotechnology News.
Apa Reaksi Anda?
 Suka
0
Suka
0
 Kurang Suka
0
Kurang Suka
0
 Setuju
0
Setuju
0
 Tidak Setuju
0
Tidak Setuju
0
 Bagus
0
Bagus
0
 Berguna
0
Berguna
0
 Hebat
0
Hebat
0